
 Geodiversitas
37 (3) - Pages 325-344
Geodiversitas
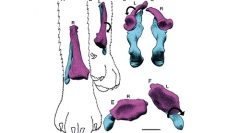
37 (3) - Pages 325-344In response to increased limb bone loads many tetrapod clades have converged upon similar adaptations to reinforce the elbow joint by reducing independent movements of the forearm bones. However prior studies have not examined how these changes occurred phylogenetically or functionally, such as during the transition from prehensile forelimbs in dinosaurs to gliding/flapping flight in bird wings. Here, a functional analysis of forearm bone mobility in extant archosaurs shows that crossing and uncrossing of the radius and ulna can be forced in alligators via a passive gliding mechanism recently described in lacertilians, while birds are adapted to inhibit this motion. A comparison of these findings with a sample of extinct quadrupedal archosaur forearms strongly suggests that, due to the highly conserved morphology of tetrapod forearms in general, the lacertilian mechanism broadly describes the plesiomorphic mechanism via which tetrapod forearm bones passively cross in response to locomotor-induced torsion. Bipedal dinosaurs retained adaptations for this passive mechanism, which indicates that they were unable to perform active long-axis rotations to align their semi-pronated, misaligned forearm joints. By contrast, analogous to birds and pterosaurs, quadrupedal dinosaurs evolved immobilizing adaptations to reduce or prohibit independent movements of the radius and ulna. Notably, the elbow joints of Archaeopteryx von Meyer, 1861 and dromaeosaurs are bird-like. This information, coupled with a lack of non-aerial adaptations for increased limb bone loads, strongly suggests that the forearms of deinonychosaurs were adapted to resist the bending and torsional stresses incurred by leading edge air streams during gliding and/or flapping.
range of motion, extant phylogenetic bracket, radial morphology, ulnar morphology, independent flexion/extension, evolution of flight
